
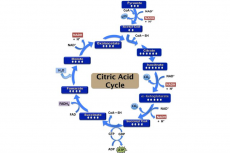
A reminder: Acetyl-CoA, citrate, iso-citrate, alpha-ketogluterate, succinate, fumerate, malate and oxaloacetate are all intermediates that are regenerated during each turn of the cycle. Additional amounts of intermediate added can be retained within the cycle due to one intermediate being converted into another, thus the amount of intermediate increases. Withdrawal of intermediates for biosynthesis has the potential to cause damage. For example, if oxaloacetate is scarce, the cycle will stop.
Therefore, intermediates must be replenished, this is achieved by anaplerotic reactions. The removal of an intermediate has a cataplerotic effect. Anaplerosis is the act of replenishing intermediates that have been extracted for biosynthesis (cataplerotic reactions.) Throughout the duration of the cycle, anaplerotic and cataplerotic reactions increase and decrease the fate of oxaloacetate availability for fusion with acetyl-CoA to form citrate. Thus, if oxaloacetate availability is limited, ATP production rate is decreased, thus less obtainable ATP in the cell. In the liver, carboxylation of pyruvate (using pyruvate carboxylase) occurs to produce oxaloacetate, replenishing the intermediate.
When proteins are broken down into amino acids by proteases, their carbon skeletons may enter the citric acid cycle as intermediates, such as alpha-ketogluterate which comes from glutamine (anaplerotic effect). Acetyl-CoA can come from leucine and other amino acids. The acetyl-CoA can be burned to carbon dioxide and water or used to form ketone bodies. These amino acids are described as ‘ketogenic.’ Glucogenic amino acids are those that can be converted into glucose through gluconeogenesis. Those that enter the citric acid cycle as intermediates can only be cataplerotically removed through the gluconeogenenesis pathway via malate. Deaminated amino acids (such as alanine, glycine and serine) can be converted into pyruvate. Pyruvate can either proceed to the link reaction, to form acetyl-CoA, to be disposed of as carbon dioxide and water in the cycle or can be converted into oxaloacetate through carboxylation.
Triglycerides are hydrolysed to produce fatty acids and glycerol in fat catabolism. The liver can convert glycerol into glucose through gluconeogenesis using dihydroxyacetone phosphate and glyceraldehyde-3-phosphate. The process of breaking down fatty acids (beta oxidation) results in mitochondrial acetyl- CoA production which can then be used in the citric acid cycle. The intermediate succinyl-CoA, which can contribute to the anaplerotic effect in the cycle, can be converted from propionyl- CoA. Propionly-CoA is a product of beta oxidation from fatty acids with an odd number of methylene bridges.
Thus, intermediates of the citric acid cycle serve as substrates for anabolic biosynthetic processes. Acetyl-CoA can be used for the production of cholesterol and fatty acids. Synthesis of steroid hormones, vitamin D and bile salts can occur through the use of cholesterol. However, acetyl-CoA cannot be transported out of the mitochondrion, instead it must be obtained from citrate. Citrate is removed from the cycle and is transferred to the cytosol where it is cleaved by ATP citrate lyase into acetyl-CoA and oxaloacetate. Acetyl-CoA is used whilst oxaloacetate is returned to the cycle as malate where it continues participating in the Krebs cycle.
Mitochondrial oxaloacetate is reduced to malate during gluceoneogenesis, then malate is transported into the cytosol where it is oxidised back into oxaloacetate. Using phosphoenolpyruvate carboxykinase, oxaloacetate is then decarboxylated to phosphoenolpyruvate. Phosphoenolpyruvate limits the conversion of gluconeogenic precursors, such as lactate, into glucose by the liver.
image- https://www.thoughtco.com/citric-acid-cycle-373397

0 Comment:
Be the first one to comment on this article.